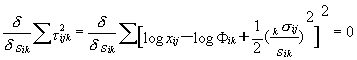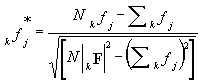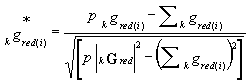and k = 1 |
(3b) |
Diversity of ecological groups of species in cultural forests of South Bohemia
Karel Matějka
Published in Ekológia (Bratislava), Vol. 12 (1993), No. 3, 299-316.
Abstract
Species diversity is important, nevertheless not the sole diversity, which could be studied in ecosystems. Also the diversity of ecological groups of species (hereinafter called ecological diversity) is of importance. A functional relation between stability and ecological diversity is supposed.
To estimate the ecological diversity, a new bioindication method (IDGAI) has been used, based on the system of eco-indices (for used system see Ellenberg, 1974). The relation of species diversity, number of species, and indices of ecological diversity related to six environmental factors (light conditions, temperature, degree of oceanity/continentality, soil moisture, soil reaction, nitrogen activity) has been studied within the cultural forests of South Bohemia.
Introduction
The article attempts to answer following questions:
Before the analysis of the problematics, some approaches and terms have to be explained:
Each species has an optimum and an amplitude of abundance at the gradient of each ecological factor. In this article we are speaking about the ecological optimum. The question of a normal-distribution-shape of the species response function is often discussed. In fact it is a pseudoproblem - the normality can be reached by an acceptable transformation of each variable describing the ecological factor level.
The definition of ecological groups of species is another problem. Having in account continuity of the species response on ecological factor level, it is obvious that strictly limited groups of species cannot exist. Similarly to the conception of the syntaxonomical unit as an arbitrary type, we can speak about certain types of behaviour of species, and those inclining in their real behaviour to one type can be included into one ecologic group.
The problem of representation of different ecological and plant coenological groups of species in different types of phytocoenose is mentioned in many papers. Nevertheless the results can be quantified with problems, due to a not very exact definition of groups of species, and different levels of definition of those groups along the gradient of environment.
Currently, the great amount of information of different plant species is available, nevertheless only few systems present complete knowledge of the behaviour of all (better to say of most) species in certain large region. The Ellenberg's system of eco-indices (Ellenberg, 1974, 1979) (although often critizied, the legitimacy left apart), is one of them.
The use of Ellenberg's eco-indices as well as other similar tables (Landolt, 1974; Zólyomi et al., 1967, etc.) was described in many papers (for instance Jurko, 1981, 1983a, b, 1984, 1985; Jurko, Kubíček, 1979; Jurko, Kubíček, Šomšák, 1981; Jurko, Kontriš, 1981, 1982; Rambousková, 1981, 1982, 1984 from Czechoslovakia; an interesting example was given by Reif, Teckelmann, Schulze, 1985). Different authors have a very distinct meaning, from an uncritical application of this method (several papers from Jurko above cited and others) to discounting for any using. Discussion to the problem linking with ecological indication has brought Mucina (1985) and Jurko (1986). There are two principles of a reasonable approach to this problematic:
Hereinafter the second approach will be kept. It is based on a premise that ecological optima and value of eco-indices have equal or very similar trend (it means the arrangement of species according to eco-indices is little different from arrangement according to ecological optima). An often mentioned problem of a no-Gaussian response of some species along gradients (as bimodal distribution) is solved by not involving such species into group of indicators.
Assuming results of ordination modes using previously obtained (a priori) information about species' ecological characteristics, it would be better to compare them with the classical methods of indirect gradient analysis. After evaluating some uses of bioindication approaches (Clausman, 1980; Ter Braak, Gremmen, 1987), a new method, called In-Direct Gradient Analysis with apriori Information (IDGAI) was developed for this purpose. It was developed at the Institute of Soil Biology of the Czech Academy of Sciences in Ceské Budejovice by the author of this paper. An application of it for grass vegetation see in Matějka (1996), the method description is presented in the following text.
To specify the definition of species diversity is not necessary, but it is important to explain, what does the term "diversity of ecological groups of species" (i.e. ecological diversity) mean. In the context of ecological groups of species, the question is the diversity of the requirements of species on the level of environmental factor. A higher diversity consists in a more differentiated ecological optimum of certain species and their closer amplitudes (i.e. more specified demands).
Data collection and description of forest groups
The material from plant coenological study realized in 1982-1989 in the phytogeographic districts of 37h (Prachatické Předšumaví), 37i (Chvalšinské Předšumaví) and 38 (Českobudějovická pánev) with adjacent regions (37g, 37j, 37l; see Skalický in Hejný et Slavík, 1988) has been used. Studied forests grow at localities the altitudes of which are from 370 to 1100 m. Average year temperature varies from 4.5 to 8 °C and the year total of precipitations varies from 570 to 800 mm (Vesecký et al., 1958).
The total of 63 relevés has been classified. The sample area varied from 200 to 400 m2. For the purpose of our work, data of herb layer of forests were analysed. Using the divisive classification procedure TWINSPAN (Hill, 1979), following four classification groups of phytocenoses have been stated:
The IDGAI method description
The used designation: The primary data matrix X=[xij], 0<i<P and 0<j<N, where P is the total number of species in the data set, N is the total number of samples, xij
is the value of quantitative contribution of i-th species in j-th sample (usually species cover). Let us assume that Σxij=1 (over species for each sample). The term 'relevé' is reserved for the list of species occurred in a sample (at some plot of certain area) with given values of species significance. The vector of primary (in some table contained) ecological characteristics (eco-indices) of species is a column vector 0G, 0G=(0g1,0g2,...,0gP)T (T indicate the transposing).In the first time, the vector of primary ecological characteristics of samples is computed as the weighted arithmetical mean of the species eco-indices
0F = 0GT.X = (0f1,0f2,...,0fN) |
(1) |
In reality, this mean is given over p species with known eco-indices only (p≤P) [kGred or Xred(k) arise drom kG or X by omitting all rows for which kgi is not known and we normalize Xred(k) to the sum equal to one for every column]
0F = 0GredT.Xred(0) |
(2) |
One can order of samples along the hypothetical gradient of 0fi. Let we assume the normal distribution of each species along such a gradient in the form
xij = Φik exp{(1/2)[(kσij)/sik]2} + τijk |
(3) |
where
Φik = Pik / [sik √(2π)] |
(3a) |
and
kσij = (k-1)fj-kgi
by
and k = 1 |
(3b) |
Under the condition that
|
(4) |
acquire the least value, it is with satisfying the conditions
|
(4a) |
and
|
(4b) |
(we compute sums for n no-zero value of xij only, n ≤ N [We may fill a condition that the sum is calculated for xij > x* only; x* ≥ 0 is the earlier fixed real value]), one can estimate the parameters kgi, sik
and Φik (or Pik). In this model we give the follow names to the parameters: kgi - eco-index of the i-th species, kfj eco-index of the j-th relevé, sik the measure of the i-th species tolerance, Pik the eco-index weight coefficient of the i-th species.Average of such k-th iteration of eco-indices of species is allowed to write as
kG = X . (k-1)FT |
(5) |
The parameters sik and Ŕik are estimated by solving the two equations (4a) and (4b), that is by solving the matrix equation
|
(6) |
The items of the matrix A (dimension 2*2) and the column vector B (dimension 2*1) are
|
|
|
|
|
|
(the condition for summing as in eq. (4a) and (4b)).
If the condition 4 (and eq. (6)) have not a solution (for sik is from R+) or sik > smax (for fixed smax>0) let us declare i-th species be indifferent (in k-th iteration) to the environmental factor studied, consequently kgi will not be defined (see eq. (5)).
Based on the computing the 1G vector (in the first iteration or the kG vector in the k-th iteration) or 1Gred as a more real case may be, the computing of 1F (or kF) is possible similarly to the equation (2):
kF = kGredT . Xred(k) |
(7) |
(this way is named 1st variant of IDGAI). The use of knowledge of the indication strength of species (eco-index weight coefficient Pik) seems to be more suitable. For this case, we would use the equation
|
(8) |
(summing for i in the case the kgi value is set up) (the 2nd variant of IDGAI).
The possibility of iteration of this algorithm has been suggested earlier. It is carry out continuing by eq. (3) for k increased of one.
Concluding, the parameters differing several ways of IDGAI are as following:
1. The variant of IDGAI (selection between eq. (7) and (8));
2. The level of significant species representation x*;
3. The upper limit of standard deviation, smax (it is admissible smax=+∞ by the 2nd variant).
Regarding to decrease of standard deviation of kfj (kgi) for increasing iteration degree k, the indices normalization is acceptable:
|
(9a) |
|
(9b) |
The last choice is
4. The normalization of the indices (eq. (9a) and (9b)).
Methods
The degree of combined scale for abundance and dominance of species of herb layer from the registrated relevés was transformed into the percentage of average covering. This was standardized to the sum of significance of all species equal to the total cover of herb layer. The known values of eco-indices for light conditions, temperature, level of oceanity/continentality, moisture, soil reaction and nitrogen activity were assigned to all species. Before next computation, values of the average eco-indices of the relevé were transformed into real scale using the function
![]()
where M is the maximum permissible value of respective eco-index (M = 9; M = 12 for soil moisture only). The data were processed by the method IDGAI. Similarly as the average eco-index of relevé computed using the weight (equal to the ratio of the real species cover and index P, see eq. (8)), also the standard deviation of average eco-index was computed. This value is considered to be the measure of ecological diversity.
Species diversity and their components were described by three basic indices (Shannon-Wiener's index of species diversity H', total number of species S and equitability e=H'/log2S). The relation among all the values described has been studied using current statistical methods.
Results
Indices of the first iteration of the procedure IDGAI (the second variant using eq. (8)), with following parameters, were used:
The minimum, maximum and average of eco-indices of relevés for different environmental factors (the number of relevés with assignated eco-indices is presented in brackets):
light conditions |
-0.837 |
0.864 |
0.152 |
(19) |
temperature |
-0.229 |
-0.005 |
-0.092 |
(4) |
oceanity/continentality |
-1.182 |
0.125 |
-0.564 |
(28) |
soil moisture |
-1.472 |
0.411 |
-0.305 |
(51) |
soil reaction |
-3.490 |
1.196 |
-0.500 |
(49) |
nitrogen activity |
-2.140 |
1.644 |
-0.090 |
(50) |
The fact, that the first three indices describing global characteristics of the environment are of very limited descriptive capacity and utility, and that the last three describing soil characteristics are much more suitable, is evident. Therefore in the following text an attention will be paid to those three later ones. The survay of the computed eco-indices and of the indices of diversity is presented in Table 1. Some species - indicators are listed in Table 2.
The correlation between the index of species diversity (Shannon-Wiener index H'), their compounds (number of species S and equitability e) and
ecological diversity related to different environmental factors have been studied. At the level α = 0.05, the correlation between ecological diversity related to light conditions and ecological diversity related to the level of oceanity/continentality (r = 0.620, n = 12) is significant. At the level α = 0.005 the ecological diversity in relation to moisture and diversity related to soil reaction (r = 0.537, n = 45) is significant, too.The distribution of relevés in the space of three axis of ecological diversity in relation to soil reaction, to nitrogen activity and to moisture (Fig. 1) shows the existence of three groups of plant coenoses. The group of low (same as the group of high) levels of these ecological diversity indices are not constituated any specific types of plant communities. The low ecological diversity is connected mainly with plant coenoses with a low number of species, the high diversity can be observed in coenoses with medial or higher number of species. The fact, that the relation between ecological diversity related to soil reaction and the one related to nitrogen activity is not significant, is also of interest (Fig.2). Next two relations are presented at the Fig. 3.
The correlation between indices of species diversity and indices of ecological diversity is not significant, nevertheless in data obtained had been observed some symptoms of the positive correlation (with the exclusion of the ecological diversity related to oceanity/continentality, where negative values of coefficients of correlation to indices H', e, S were measured; all values were statistically nonsignificant). It is not possible to say, which of three indices of species diversity (H', e, S) is more affecting the value of the indices of ecological diversity (there is a different index for each factor studied).
Following characteristics are typical for ecological diversity of groups of forest phytocenoses:
Attention is to be paid also to phytocenoses of high values of ecological diversity indices (see Fig. 4), where higher stability in conditions of changes of environmental factor is supposed. In the history of those phytocenoses, a higher fluctuation of the level of this environmental factor can be supposed. Whether it is really a higher ecological diversity must be considered always in relation to the indicated level of the factor: in a limit level of environmental factor, also a relatively lower value of the index of ecological diversity in comparison with phytocenoses of non-limit level of this environmental factor can be considered to increas (see Fig. 4A). In phytocenoses of a limit level of environmental factor more frequent occurrence of increased ecological diversity can be expected (see Figs. 4C and 4D).
Discussion and conclusion
Attention is to be paid to the relation of diversity and stability of phytocenoses. Species diversity was mistaking for ecological diversity which resulted in many errors. Similar approach could be useful also for the solution of the relation between diversity and productivity - in this case the study of the niche diversity would be more pertinent. Nevertheless, it is supposed that there can be a certain relation between diversity of niche and diversity of ecological groups of species.
It is probable that the lower diversity of ecological groups of species causes the lower stability of coenose (without consideration of species diversity).
In that sense also the effect of liming of acid forest soil with purpouse of compensation of the influence of acid rains can be contemplated. In naturally acid soil of mountain spruce forests different edafon species are prevailing, adapted to low values of soil reaction. Many of them are intolerant to lime added and thus eliminated by liming. Only species of wide amplitude of the response on changes of the factor "soil reaction" are surviving. Thus not only species diversity, but also diversity of ecological groups of species is decreased. It is a reason for the decrease of stability of the community.
In such a way, relations among different terms used in current ecology of phytocenoses can be mentioned: definition of coenose, continuum, coenological units, ecotones, spatial organization of sub-coenological level, species diversity, diversity of ecological groups of species, niche diversity, diversity of units of sub-coenological level, stability (resistance, resilience, persistence), stress (acute, chronic), environmental factors (continuity in space, discrete or continuous levels) and others.
Other terms related could be:
References
Clausman, P.H.M.A., 1980: Ecologische interpretatie van vegetatieopnamen m.b.v. een computer. WLO-Meded., 7, p.92-98.
Ellenberg, H., 1974: Zeigerwerte der Gefässpflanzen Mitteleuropas. Scripta Geobotanica, Vol. 9, Göttingen, 97 pp [2nd Ed., 1979].
Hill, M.O., 1973: Reciprocal averaging: an eigenvector method of ordination. Journ. Ecol., 61, p.237-249.
Hill, M.O., 1979: TWINSPAN - a FORTRAN program for arranging multivariate data in an ordered two way table by classification of individuals and attributes. Cornell Univ. Ithaca, 48 pp.
Jurko, A., 1981: Ökospectrum und Stabilität der Waldgesellschaften. Biológia (Bratislava), 36, p.497-507.
Jurko, A., 1983a: Survey of habitat indication by plants and leaf categories in forest ecosystem types of the Little Carpathian mountains. Ekológia (ČSSR), 2, p.49-74.
Jurko, A., 1983b: Trampling effect on species diversity and leaf characteristics of vegetation in the High Tatras Mts. Ekológia(ČSSR), 2, p.281-293.
Jurko, A., 1984: Ökologische Indikationswerte der Pflanzengesellschaften der Ostslowakischen Tiefebene. Ekológia (ČSSR), 3, p.413-425.
Jurko, A., 1985: A contribution to ecological diversity of some plant communities. Ekológia (ČSSR), 4, p.399-406.
Jurko, A., 1986: Notes to the discussion about the usefulness of Ellenberg's indication values (in Slovak). Biólogia (Bratislava), 41, p.91-100.
Jurko, A., Kontriš, J., 1981: Ecological profiles of forest biocenoses in the Little Carpathians. Acta Ecol., 9/24, p.7-63.
Jurko, A., Kontriš, J., 1982: Kulturelle Nadelforstgesellschaften in den Kleinen Karpaten. Biológia (Bratislava), 37, p.909-918.
Jurko, A., Kubíček, F., 1979: Ökologische Analyse der Waldgesellschaften des Silica-Plateaus. Biologické Práce, 25/2, p.91-201.
Jurko, A., Kubíček, F., Šomšák, L., 1981: Ecological and production characteristics of the undergrowth of mountain forest. Folia Geobot. Phytotax., 16, p.153-179.
Klimeš, L., 1987: Exploitation of tabulated indicator values in the gradient analysis of vegetation (in Czech). Preslia, Praha, 59, p.15-24.
Landolt, E., 1974: Ökologische Zeigerwerte zur Schweizer Flora. Veröff. Geobot. Inst. ETH, Stift. Rübel, Vol. 64, Zürich.
Matějka, K., 1993: Ecology and coenology of grass species in meadows and herb communities of the south foothills of Šumava Mts. (in Czech). Zprávy Čes. Botanické Společnosti, Praha, Mater. 13, p. 71-77.
Mucina, L., 1985: To use or not to use the Ellenberg's indication values? Biológia (Bratislava), 40, p.511-516.
Neuhäusl, R., Neuhäuslová-Novotná, Z., 1979: Natural forest vegetation of the Železné hory Mts. (in Czech). Studie ČSAV, Praha, 2(1979), 203 pp.
Rambousková, H., 1981: A contribution to the application of some numerical indices in the evaluation of recreational impact on vegetation. Preslia, Praha, 53, p.147-158.
Rambousková, H., 1982: Recreation-induced changes in natural and seminatural vegetation with a special respect to camping activites. Ekológia (ČSSR), 1, p.313-330.
Rambousková, H., 1984: Some notes on forest ruderalization. Preslia, Praha, 56, p.117-123.
Reif, A., Teckelmann, M., Schulze, E.D., 1985: Die Standortamplitude der Grossen Brennessel (Urtica dioica L.) - eine Auswertung vegetationkundlicher Aufnahmen auf der Grundlage der Ellenbergschen Zeigerwerte. Flora, Jena, 176, p.365-382.
Ter Braak, C.J.F., Gremmen, N.J.M., 1987: Ecological amplitudes of plant species and the internal consistency of Ellenberg's indicator values for moisture. Vegetatio, 69, p.79-87.
Vesecky, A. et al., 1958: Climate atlas of Czechoslovakia (in Czech). Praha.
Zlatník, A., 1976: Forest phytocenology (in Czech). Praha.
Zólyomi, B. et al., 1967: Einreihung von 1400 Arten der ungarischen Flora in ökologische Gruppen nach TWR-Zahlen. Fragm. Bot. Mus. Hist. Nat. Hung., 4, p.101-142.
See also
Back to the main page IDS
© Karel Matějka - IDS (2003)